
Tu manges trois fois par jour. Tu ne le penses pas comme ça, mais à chaque repas, tu envoies un signal moléculaire à tes cellules.
Pas une métaphore. Un signal chimique, traçable, mesurable, qui décide concrètement quels gènes seront lus dans tes cellules et lesquels resteront silencieux.
Le mécanisme central qui fait ce travail s’appelle la méthylation de l’ADN. C’est l’un des principaux interrupteurs épigénétiques connus. Et il est massivement piloté par ce que tu apportes — ou n’apportes pas — à ton corps via ton assiette.
On va être clair. Tu n’es pas le pantin de tes gènes. Mais tu n’es pas non plus la victime de ton alimentation. Le lien entre méthylation ADN et alimentation est un terrain solide : ce qui est établi est solide, ce qui reste ouvert mérite d’être posé honnêtement, et ce qui est extrapolé abusivement par certains discours bien-être doit être nommé pour ce que c’est.
Si tu as déjà lu ton environnement reprogramme tes cellules : ce que l’épigénétique change à tout, tu connais le cadre général. Cet article descend d’un cran : on entre dans le mécanisme moléculaire concret. Comment, exactement, une vitamine B9 absorbée à midi modifie-t-elle l’expression d’un gène le soir ?
Qu’est-ce que la méthylation de l’ADN, concrètement
Avant les études, il faut comprendre la mécanique. Sinon tout le reste glisse.
Une marque chimique posée sur tes gènes
Ton ADN est une longue chaîne de quatre bases : adénine (A), thymine (T), guanine (G), cytosine (C). C’est l’alphabet de ton patrimoine génétique. Cet alphabet ne change pas (sauf mutations rares).
La méthylation consiste à coller un petit groupe chimique — un groupe méthyle (-CH₃, un carbone et trois hydrogènes) — sur certaines cytosines de l’ADN. Précisément sur les cytosines qui sont suivies d’une guanine. On parle alors de sites CpG.
Cette marque ne modifie pas la séquence de l’ADN. Elle se pose dessus, comme un autocollant chimique. Mais elle change tout sur le plan fonctionnel : un gène fortement méthylé sur sa zone de démarrage (le promoteur) est généralement silencieux. Il ne sera pas transcrit, donc pas traduit en protéine. Comme si on avait collé un ruban “ne pas ouvrir” sur ce livre de la bibliothèque.
À l’inverse, un gène faiblement méthylé sur son promoteur est généralement actif. Il fonctionne.
Pourquoi ça change tout
Toutes les cellules de ton corps contiennent le même ADN. Le foie, les neurones, les cellules musculaires, tes globules blancs : même bibliothèque. Et pourtant, un neurone ne fait pas le travail d’un hépatocyte.
Ce qui les différencie, c’est précisément quels gènes sont activés et lesquels sont étouffés. La méthylation est l’un des principaux mécanismes qui décident ça.
Et la méthylation n’est pas figée. Elle évolue tout au long de ta vie en fonction de ton environnement, de ton alimentation, de ton stress, de tes expositions. C’est ce qui fait de l’épigénétique un terrain bien plus dynamique que la génétique.
Concrètement : si demain tu changes ton alimentation, ton cycle de méthylation se modifie en quelques semaines. Pas tous les sites, pas tous les gènes — mais une fraction significative. Et ça se mesure en laboratoire.
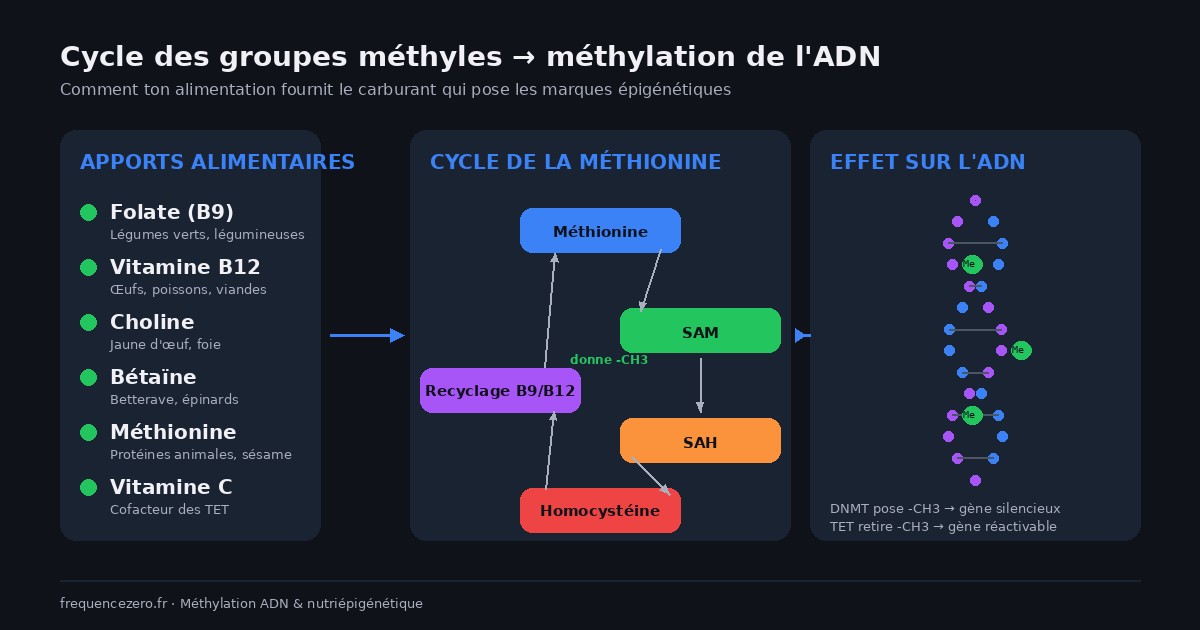
Le mécanisme moléculaire : le cycle des groupes méthyles
C’est ici que ton assiette entre dans la danse. Parce que le groupe méthyle qui finit collé à ton ADN, il vient de quelque part. Et ce quelque part, c’est ton métabolisme — qui dépend des nutriments que tu absorbes.
SAM, le donneur universel de méthyles
La molécule centrale s’appelle S-adénosylméthionine, abrégée en SAM. C’est le donneur universel de groupes méthyles dans ton corps. Pas seulement pour l’ADN — aussi pour la synthèse de neurotransmetteurs (dopamine, mélatonine), pour la détoxification hépatique, pour la production de phosphatidylcholine. La SAM, c’est une devise interne très demandée.
La SAM se forme à partir de la méthionine, un acide aminé essentiel. Une fois qu’elle a donné son groupe méthyle, elle se transforme en S-adénosylhomocystéine (SAH), puis en homocystéine. À ce stade, deux options : recycler l’homocystéine en méthionine (ce qui demande du folate et de la vitamine B12), ou l’envoyer dans la voie de transsulfuration (qui produit du glutathion, un antioxydant majeur).
Tout ce circuit s’appelle le cycle de la méthionine, ou cycle des groupes méthyles. Il tourne 24h/24 dans toutes tes cellules. Et il est entièrement dépendant de tes apports alimentaires.
Les enzymes DNMT, qui posent les marques
Les enzymes responsables de la méthylation de l’ADN s’appellent les DNA méthyltransférases, ou DNMT. Trois principales : DNMT1, DNMT3A, DNMT3B. Elles vont chercher la SAM, prélèvent le groupe méthyle, et le transfèrent sur les cytosines des sites CpG.
DNMT1 est l’enzyme de maintenance : à chaque division cellulaire, elle copie le motif de méthylation parental sur l’ADN nouvellement synthétisé. Sans elle, l’information épigénétique se perdrait à chaque mitose.
DNMT3A et 3B sont les enzymes de novo : elles posent de nouvelles marques de méthylation, en réponse à des signaux internes ou environnementaux. Ce sont elles qui répondent en partie aux signaux nutritionnels.
Les enzymes TET, qui retirent les marques
La méthylation n’est pas une condamnation à perpétuité. Une famille d’enzymes appelée TET (TET1, TET2, TET3) peut retirer ou modifier les marques de méthylation. Elles oxydent le groupe méthyle en plusieurs étapes, jusqu’à le faire éliminer par le système de réparation de l’ADN.
Ces enzymes TET dépendent de cofacteurs précis : du fer, du 2-oxoglutarate, et de la vitamine C. Une carence en vitamine C, par exemple, ralentit la déméthylation. Ce n’est pas un détail : ça veut dire que ce que tu manges agit aussi sur la capacité de ton corps à effacer des marques.
Le vrai sujet, c’est celui-ci : la méthylation de ton ADN n’est pas un état figé, c’est un équilibre dynamique entre pose et effacement, entièrement nourri par ton métabolisme.
Méthylation ADN alimentation : les nutriments qui pilotent l’expression de tes gènes
On arrive au cœur. Quels nutriments précis font tourner ce cycle, et qu’est-ce qui se passe quand ils manquent ?
Le folate (vitamine B9), le donneur initial
Le folate est probablement le nutriment le plus étudié de la nutriépigénétique. Il fournit le carbone qui finira sur la SAM, puis sur l’ADN. Une carence en folate diminue le pool de SAM disponible, ce qui réduit la capacité de méthylation de tes cellules.
Tu trouves le folate dans les légumes à feuilles vertes (épinards, mâche, roquette, cresson), les légumineuses (lentilles, pois chiches, haricots blancs), le foie, et les œufs. La forme synthétique (acide folique) ajoutée à certains aliments enrichis est moins efficace chez environ 30 à 40 % des personnes porteuses du polymorphisme MTHFR C677T, qui ralentit la conversion de l’acide folique en folate actif. Pour ces personnes, la forme méthylfolate (5-MTHF) est mieux assimilée.
L’étude néerlandaise de la famine de 1944-1945, déjà détaillée dans l’article fondateur sur l’épigénétique, repose en grande partie sur ce mécanisme : la sous-nutrition maternelle, particulièrement en folate, a laissé des marques de méthylation modifiées chez les enfants conçus durant cette période — des marques toujours détectables 60 ans plus tard.
La vitamine B12 (cobalamine)
La B12 est cofacteur de l’enzyme qui recycle l’homocystéine en méthionine. Sans elle, le cycle s’enraye. Une carence prolongée en B12 augmente l’homocystéinémie, marqueur indirect d’un cycle de méthylation grippé, et associé à un risque cardiovasculaire et neurodégénératif.
La B12 est exclusivement d’origine animale (viandes, poissons, œufs, produits laitiers). Les régimes végétaliens stricts non supplémentés mènent à une carence quasi inévitable à 3-5 ans. Si tu suis ce type de régime, supplémente. Ce n’est pas une opinion, c’est un fait métabolique.
La choline et la bétaïne
La choline est un nutriment essentiel souvent oublié. Elle peut être convertie en bétaïne, qui sert de donneur alternatif de méthyles via une voie indépendante du folate. Quand le folate manque, la voie choline-bétaïne prend le relais — partiellement.
Sources principales : jaunes d’œufs, foie, poissons gras, soja, brocolis. Une grande partie de la population française est en apport sub-optimal en choline, surtout les femmes et les personnes qui évitent les œufs. Les besoins augmentent pendant la grossesse — c’est probablement l’un des nutriments les plus importants pour le développement épigénétique du fœtus, avec le folate.
La méthionine
La méthionine elle-même est apportée par les protéines animales (viande, poisson, œufs, produits laitiers) et, en moindre quantité, par certaines protéines végétales (graines de sésame, noix du Brésil, soja). Un apport très bas réduit le substrat de la SAM.
Mais attention : un apport très élevé en méthionine n’est pas non plus optimal. Les régimes hyperprotéinés chroniques ont été associés, dans plusieurs études animales, à des modifications épigénétiques peu favorables. Comme pour beaucoup de choses dans le métabolisme : la dose fait l’effet.
Les polyphénols : resvératrol, EGCG, curcumine, sulforaphane
Plusieurs molécules végétales agissent directement sur les enzymes DNMT et TET. Le resvératrol (peau de raisin), l’EGCG (thé vert), la curcumine (curcuma), et le sulforaphane (brocoli, choux) ont tous démontré, in vitro et chez l’animal, des effets de modulation de la méthylation. Certains inhibent les DNMT, d’autres activent les TET, d’autres encore agissent sur les histones (autre couche épigénétique).
On ne va pas se mentir : la traduction de ces effets in vitro à l’humain est délicate. Les concentrations utilisées en laboratoire sont souvent très supérieures à ce qu’une alimentation normale fournit, et la biodisponibilité de la curcumine, par exemple, est notoirement faible. Manger varié et coloré reste la stratégie la mieux soutenue par les données. Acheter une dose géante de curcumine en gélules pour “réguler son épigénome” est, à ce stade, une projection plus qu’une certitude.

Nutriépigénétique : ce que les études montrent vraiment chez l’humain
Je vais être clair : la majorité des données solides en nutriépigénétique vient d’études animales et d’études d’observation chez l’humain. Les essais randomisés de longue durée sont rares, parce qu’ils sont longs et coûteux à mener proprement. Voilà ce qui est solide à ce jour.
La famine hollandaise et ses descendants
L’étude Dutch Hunger Winter (Heijmans et al., 2008 ; Lumey et al., 2007) reste l’une des démonstrations les plus parlantes. Les enfants conçus pendant la famine de l’hiver 1944-1945 présentent, six décennies plus tard, une hypométhylation persistante du gène IGF2 (insulin-like growth factor 2). Et une incidence supérieure de maladies cardiovasculaires, de diabète et d’obésité.
Le mécanisme est plausible et reproduit chez l’animal. C’est l’une des preuves les plus robustes que la nutrition pendant des fenêtres critiques (grossesse, petite enfance) laisse une trace épigénétique durable.
Les abeilles et la gelée royale
Chez l’abeille, deux femelles génétiquement identiques peuvent devenir l’une ouvrière (stérile, durée de vie de quelques semaines), l’autre reine (fertile, durée de vie de plusieurs années). La seule différence : la larve future-reine est nourrie exclusivement à la gelée royale.
La gelée royale contient une molécule appelée royalactine, qui modifie la méthylation de l’ADN de la larve via l’inhibition d’une DNMT. Le résultat : un destin biologique radicalement différent à partir d’un signal alimentaire. Kucharski et al. ont démontré ce mécanisme en 2008 dans Science.
C’est l’une des illustrations les plus nettes de ce que la nutrition peut faire à l’expression génétique. Évidemment, on ne peut pas extrapoler tel quel à l’humain. Mais le principe — un nutriment qui modifie une enzyme épigénétique, qui modifie la trajectoire de développement — est universel chez les eucaryotes.
Les études d’intervention humaine
Plusieurs essais ont mesuré l’effet d’interventions nutritionnelles sur la méthylation chez l’adulte. Quelques résultats robustes :
Une supplémentation en folate chez l’adulte modifie la méthylation de plusieurs gènes en quelques mois (Crider et al., 2012). L’amplitude est modérée, mais reproductible.
Le régime méditerranéen, dans l’étude PREDIMED (Arpón et al., 2017), s’accompagne d’une modification du profil de méthylation associée à une réduction du risque cardiovasculaire. La signature épigénétique observée est cohérente avec les bénéfices cliniques.
Une restriction calorique modérée (sans dénutrition) modifie la méthylation des gènes liés à l’inflammation et au métabolisme. C’est l’un des mécanismes proposés pour expliquer pourquoi les modèles animaux soumis à restriction calorique vivent plus longtemps.
Le jeûne intermittent, plus controversé sur le plan épigénétique humain, semble agir partiellement par ce levier, mais les données chez l’humain sont encore limitées.
Ce qui reste ouvert dans la recherche sur la méthylation ADN
Honnêtement, plusieurs zones du dossier sont loin d’être tranchées.
La causalité directe entre une intervention précise et un effet clinique précis reste difficile à établir. On observe des corrélations entre profils alimentaires et profils de méthylation. On observe des corrélations entre profils de méthylation et issues de santé. Le lien causal complet, étape par étape, sur des décennies, n’est documenté que pour quelques gènes bien identifiés.
Les effets transgénérationnels chez l’humain sont suggérés par quelques études (descendants de la famine hollandaise, descendants de survivants de la Shoah étudiés par Yehuda et al., 2016), mais leur ampleur réelle et leur mécanisme restent débattus. Une partie de la communauté scientifique appelle à la prudence sur l’interprétation : ce qui est solide chez les rongeurs ne l’est pas toujours autant chez l’humain.
Les “âges épigénétiques” (Horvath clock, GrimAge, PhenoAge) qui prétendent mesurer ton vieillissement biologique à partir de ta méthylation, sont des outils de recherche prometteurs. Ils sont aussi commercialisés à des particuliers à des prix élevés sans toujours la nuance qui s’imposerait. Leur signal est réel, leur précision individuelle reste limitée. À considérer comme une indication, pas un verdict.
L’effet des compléments isolés (resvératrol en gélules, curcumine, NMN, etc.) sur la méthylation chez l’humain en bonne santé reste largement spéculatif. Le marketing court vite, la science marche.
Comment agir concrètement sur ta méthylation ADN au quotidien
Pas de promesse magique. Mais des leviers réels, soutenus par les données, et accessibles.
D’abord, assure les apports en cofacteurs de base. Folate, B12, choline, bétaïne, vitamine C, zinc. Ce sont les briques du cycle. Une alimentation incluant légumes verts, légumineuses, œufs entiers, poissons, et légumes crucifères couvre l’essentiel. Si tu suis un régime végétal strict, supplémente en B12 — non négociable.
Ensuite, diversifie tes sources de polyphénols. Thé vert plusieurs fois par semaine, baies, choux et brocolis, herbes aromatiques, huile d’olive vierge, un peu de curcuma frais ou en cuisine grasse. La diversité des molécules apportées vaut mieux qu’une dose massive d’un seul composé en gélule.
Puis, regarde les fenêtres critiques. Les apports en folate et choline pendant la grossesse, et la qualité alimentaire dans les 1 000 premiers jours de l’enfant, sont les leviers les plus solidement étayés en termes d’impact épigénétique durable. Si tu es concerné(e), c’est là que ton effort a le plus de poids.
Enfin, n’isole pas la nutrition du reste. Le stress chronique, le sommeil, l’exposition aux toxiques et l’environnement physique influencent aussi ta méthylation. Ce qui se joue dans ton lieu de vie compte autant que ce qui se joue dans ton assiette — pour saisir cette dimension, ce que ton lieu de vie fait à ton corps est le bon point d’entrée. La cohérence du système global fait la différence, plus qu’un super-aliment isolé.
Si tu vis dans un environnement saturé de champs électromagnétiques et que tu manges parfait, tu n’es pas optimal. Si tu manges moyennement et que ton sommeil est protégé, ton lieu calme, ton stress géré, tu fais probablement déjà mieux que beaucoup. Le cycle de méthylation aime la régularité, pas la perfection.
En résumé
La méthylation de l’ADN est l’un des principaux mécanismes par lesquels ton corps décide quels gènes sont actifs et quels gènes sont silencieux. Elle dépend d’un cycle métabolique entièrement nourri par ton alimentation : folate, B12, choline, bétaïne, méthionine fournissent les groupes méthyles, et les polyphénols ainsi que la vitamine C modulent les enzymes qui les posent ou les retirent.
Ce n’est ni magique, ni anecdotique. C’est de la biochimie, mesurable, reproductible. La nutriépigénétique offre un cadre rigoureux pour comprendre pourquoi ton assiette compte au-delà des seules calories et macronutriments.
Concrètement, tu n’as pas besoin de superaliments à 80 € le pot. Tu as besoin d’apports stables en cofacteurs de base, d’une diversité de polyphénols végétaux, et de protéger les fenêtres critiques (grossesse, petite enfance). Le reste, c’est du bonus.
Et tu n’as pas besoin de surveiller ton “âge épigénétique” tous les six mois. Tu as besoin de cohérence. Sur la durée.
À retenir
La méthylation de l’ADN consiste à coller un groupe méthyle sur certaines cytosines de l’ADN, ce qui éteint ou atténue l’expression d’un gène sans modifier sa séquence. Ce mécanisme dépend du cycle de la méthionine, alimenté par le folate (B9), la B12, la choline, la bétaïne et la méthionine. Les polyphénols (thé vert, brocoli, baies, curcuma) modulent les enzymes qui posent et retirent ces marques. La famine hollandaise, la gelée royale chez l’abeille et plusieurs études d’intervention humaine confirment l’impact réel de la nutrition sur l’expression génétique. La méthylation est dynamique, partiellement réversible, et se modifie en quelques semaines à quelques mois sous l’effet d’un changement alimentaire. Les fenêtres critiques (grossesse, petite enfance) ont l’impact le plus durable. Pas de superaliment miracle : la cohérence d’ensemble compte plus qu’une molécule isolée.
Sources et lectures
- Heijmans, B.T. et al., “Persistent epigenetic differences associated with prenatal exposure to famine in humans”, Proceedings of the National Academy of Sciences (2008) — étude de référence sur les marques de méthylation persistantes chez les descendants de la famine hollandaise.
- Lumey, L.H. et al., “Cohort Profile: The Dutch Hunger Winter Families Study”, International Journal of Epidemiology (2007) — profil de la cohorte de référence en nutriépigénétique humaine.
- Kucharski, R. et al., “Nutritional Control of Reproductive Status in Honeybees via DNA Methylation”, Science (2008) — démonstration directe qu’un signal alimentaire (gelée royale) modifie la méthylation et change la trajectoire biologique.
- Crider, K.S., Yang, T.P., Berry, R.J., Bailey, L.B., “Folate and DNA Methylation: A Review of Molecular Mechanisms and the Evidence for Folate’s Role”, Advances in Nutrition (2012) — synthèse sur le folate, la SAM et les mécanismes de méthylation.
- Arpón, A. et al., “Methylome-Wide Association Study in the Context of the Mediterranean Diet (PREDIMED)”, European Journal of Nutrition (2017) — effet du régime méditerranéen sur la méthylation et le risque cardiovasculaire.
- Yehuda, R. et al., “Holocaust Exposure Induced Intergenerational Effects on FKBP5 Methylation”, Biological Psychiatry (2016) — données sur la transmission épigénétique du trauma.
- Nessa Carey, The Epigenetics Revolution (2012) — introduction grand public solide à l’épigénétique, dont la méthylation.
- Choi, S.W., Friso, S., “Epigenetics: A New Bridge between Nutrition and Health”, Advances in Nutrition (2010) — article de synthèse fondateur sur la nutriépigénétique.
- Anderson, O.S., Sant, K.E., Dolinoy, D.C., “Nutrition and epigenetics: an interplay of dietary methyl donors, one-carbon metabolism and DNA methylation”, Journal of Nutritional Biochemistry (2012) — revue détaillée des donneurs de méthyles alimentaires et du métabolisme à un carbone.
- Horvath, S., “DNA methylation age of human tissues and cell types”, Genome Biology (2013) — article fondateur des “horloges épigénétiques”, à lire pour mesurer la portée et les limites de ces outils.